РЕПАРАЦІЯ ДНК (лат. reparatio — відновлення) — процес виправлення ушкоджень ДНК, зумовлених дією різних хімічних і фізичних факторів. Передавання спадкової інформації у незмінному вигляді — найважливіша умова виживання як кожного організму, так і виду в цілому. Отже, у ході еволюції сформувалася система, яка дозволяє клітині виправляти ушкодження ДНК, викликані помилками реплікації чи впливом ушкоджувальних факторів навколишнього середовища. Усі репараційні механізми ґрунтуються на тому, що генетична інформація записана в обох ланцюгах подвійної спіралі і, якщо нуклеотидна послідовність одного ланцюга змінена, інформацію можна відновити на основі другого (комплементарного) ланцюга. Фізичні (УФ-опромінення, іонізуюча радіація) і хімічні (ксенобіотики) фактори викликають у ДНК ушкодження чотирьох типів: 1) зміни в одиничних нуклеотидах (депуринізація, дезамінування цитозину до урацилу, дезамінування аденіну до гіпоксантину та ін.); 2) зміни в парі нуклеотидів (УФ-індуковане утворення тимінових димерів, поперечні зшивки біфункціональним алкілуючим агентом); 3) розриви ланцюгів; 4) поперечні зшивки (між основами одного або різних ланцюгів, між ДНК і молекулами гістонів). Пошкоджені ділянки можуть підлягати Р., бути заміщені шляхом рекомбінації або залишатися без змін. В останньому випадку виникають мутації, які потенційно ведуть до загибелі клітини. ДНК можуть бути репаровані за допомогою спеціальних механізмів. Основний шлях репарації ДНК має три етапи. На першому змінена ділянка ушкодженого ланцюга ДНК розпізнається і видаляється за допомогою специфічних ферментів ДНК-репаруючих нуклеаз; вони здійснюють гідроліз фосфодіефірних зв’язків між пошкодженими нуклеотидами і рештою молекули ДНК, унаслідок чого в спіралі ДНК у цьому місці виникає просвіт. На другому етапі фермент ДНК-полімераза зв’язується з 3ў-кінцем ушкодженого ланцюга ДНК і заповнює цей просвіт шляхом приєднання одного нуклеотида за другим, копіюючи інформацію, яка міститься в неушкодженому матричному ланцюгу. У кінці фермент ДНК-лігаза зшиває ДНК і таким чином завершує відновлення інтактної молекули. При дії УФ-опромінення ушкодження ДНК найбільш часто виникають у клітинах бактерій та в незахищеній шкірі людини, і один із найкраще вивчених механізмів Р. — вирізання тимінового димеру, який утворюється в цих умовах. Тимінові димери виникають унаслідок взаємодії між собою в одному ланцюгу ДНК сусідніх тимінових залишків з утворенням ковалентної зшивки:
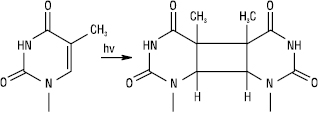
Такі димери порушують структуру ДНК і тим самим блокують її реплікацію, доки ушкодження не буде усунуте. Для видалення тимінових димерів у клітині існує два механізми: фотореактивація та ексцизійна Р. Фотореактивація відбувається при дії світла, що активує специфічний фотореактивуючий фермент — ДНК-фотолазу, який розщеплює димери на мономери і відновлює водневі зв’язки між тиміном і аденіном комплементарних ланцюгів ДНК. Ексцизійна Р. не залежить від наявності світла, тому її ще називають темновою Р. Ці два види Р. між собою дещо подібні. Відновлення нормальної структури ДНК при ексцизійній Р. здійснюється за участю комплексу ферментних систем за схемою:
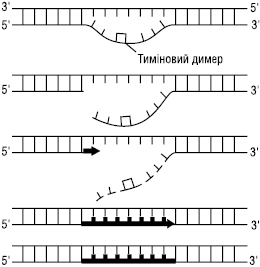
Схема. Репарація фрагмента ДНК з тиміновим димером
Спочатку діє фермент УФ-специфічна ендонуклеаза, яка знаходить місце ушкодження і розриває зв’язок поблизу від димеру з 5′-кінця. Після цього репараційна ДНК-полімераза синтезує фрагмент ділянки ДНК з неушкодженими нуклеотидами в напрямку 5′→3′, який комплементарний матричному ланцюгу. У ході цього синтезу фрагмент ланцюга, що містить тиміновий димер, видаляється. На наступному етапі ділянка, де розташований тиміновий димер, вирізається під дією 5′→3′ екзонуклеази. Нарешті, новосинтезований фрагмент з’єднується з кінцем старої частини ланцюга ДНК за участю ферменту ДНК-лігази. За допомогою темнової Р. може відбуватися виправлення більшості потенційно летальних порушень геному. Так, у бактерій вона може усувати розриви полінуклеотидних ланцюгів ДНК, викликаних дією рентгенівських променів; може видаляти зшивки пуринових основ у ДНК, викликаних дією іприту. Деякі спадкові захворювання людини пов’язані з дефектами в репарації ушкоджень ДНК, напр. пігментна ксеродерма. При цій патології шкіра пацієнтів є надзвичайно чутливою до сонячного світла, що може спричинити розвиток раку шкіри. Найбільш поширена форма пігментної ксеродерми зумовлена спадковим порушенням синтезу УФ-специфічної ендонуклеази, яка гідролізує остов ДНК поряд із тиміновим димером, що призводить до порушення всього механізму репарації ДНК. Інший шлях Р. пов’язаний з участю набору ферментів, які мають назву ДНК-глікозилази. Кожний із цих ферментів пізнає один тип змінених основ в ДНК і каталізує гідролітичне відщеплення такої основи. Вважають, що існує не менше шести типів ферментів цієї групи. Серед них ферменти, які видаляють дезамінований цитозин, дезамінований аденін, алкіловані основи різних типів та ін. Так, в ДНК можуть відбуватися зміни, зумовлені хімічною лабільністю цитозину у водневому середовищі. Залишки цитозину спонтанно, дуже повільно втрачають свою аміногрупу внаслідок гідролізу і перетворюються на залишки урацилу, який зазвичай відсутній в ДНК. Дезамінування цитозину є потенційно мутагенним, оскільки урацил, що утворився, є комплементарним аденіну, а отже, один із дочірніх ланцюгів буде містити УА-пару основ замість ЦГ-пари:
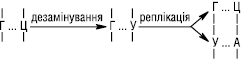
Коли новий ланцюг ДНК, що містить неправильний залишок А, у свою чергу реплікується, то в комплементарному ланцюгу на цьому місці з’явиться Т. Унаслідок цього дочірня дволанцюгова ДНК буде містити пару А–Т у тому положенні, де в материнській ДНК до ушкодження знаходилася пара Г–Ц. Ця мутація виправляється під дією репараційної системи, яка розпізнає чужерідний урацил у молекулі ДНК. Спочатку репаруючий фермент — урацил-ДНК-глікозилаза видаляє шляхом гідролізу неправильну основу, тобто урацил з ушкодженого ланцюга, утворюючи пентозо-фосфатний ланцюг без азотистої основи. Потім специфічна ендонуклеаза розпізнає цей факт і розщеплює 3′,5′-фосфодіефірний зв’язок поруч з відсутньою основою, і далі ДНК-полімераза вбудовує цитозин на місце видаленої основи. Завершує Р. ДНК-лігаза, що ковалентно зшиває розрив у ланцюгу ДНК. Аналогічним чином унаслідок дезамінування під дією азотистої кислоти, яка утворюється із нітритів і нітратів, аденін перетворюється на гіпоксантин, а гуанін — на ксантин, які виділяються із ДНК також за допомогою специфічних ферментів, після чого Р. продовжують ДНК-полімераза і ДНК-лігаза. Крім азотистої кислоти, відомі інші хімічні мутагени — інгібітори синтезу ДНК, вільні радикали, радіотоксини, аналоги пуринових і піримідинових основ, деякі антибіотики.
Р. необхідна для зберігання нативної структури генетичного матеріалу протягом усього життя організму. Тому в процесі еволюції у клітинах з’явилося багато різних механізмів, які забезпечують їх виживання і сприяють зменшенню проявів мутацій.
Гонський Я.І., Максимчук Т.П. Біохімія людини. — Тернопіль, 2001; Ленинджер А. Основы биохимии: В 3 т. — М., 1985. — Т. 3; Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: В 2 т. — М., 1993. — Т. 2.