ТРАНСКРИПЦІЯ (лат. transcriptio — переписування) — передавання інформації між різними видами нуклеїнових кислот, перша стадія реалізації генетичної інформації в клітині. Розрізняють пряму Т. (передавання інформації від ДНК до РНК) і зворотну (передавання інформації від РНК до ДНК). В еукаріотів і більшості прокаріотів відбувається пряма Т. Зворотна Т. вперше була виявлена у РНК-вмісних онкогенних вірусах, в яких вона забезпечується ферментом РНК-залежною ДНК-полімеразою (зворотною транскриптазою, або ревертазою) з утворенням ДНК-копії. В інших РНК-вмісних вірусах (грипу, кору, поліовірусу та ін.) інформація передається від РНК до РНК. На відміну від реплікації (див. Реплікація), при Т. відбувається копіювання не всієї молекули ДНК, а лише її окремих фрагментів — генів або груп генів. Тому в ДНК є сигнальні нуклеотидні послідовності, які вказують на початок та кінець ділянок молекули, що підлягають Т. (промоторні й термінальні послідовності). Гени ДНК містять інформацію про структуру всіх типів РНК і про структуру всіх білків даного виду організму. У ході Т. утворюються молекули матричної РНК (мРНК), які є матрицею для синтезу білків, а також транспортної РНК (тРНК), рибосомної РНК (рРНК) та інші види молекул РНК. Основою механізму Т., як і при реплікації, служить принцип комплементарності основ. Послідовність рибонуклеотидів у молекулі РНК комплементарна такій у дезоксирибонуклеотидів одного із ланцюгів ДНК — матричного ланцюга. У дволанцюговій ДНК, що містить багато генів, для одних генів ДНК матричним може бути один із ланцюгів, для інших генів — другий ланцюг. В еукаріотів процес відбувається в ядрі. Синтез молекул РНК починається в певних послідовностях (сайтах) ДНК — промоторах і завершується в термінувальних ділянках (сайтах термінації). Ділянка ДНК, обмежена промотором і сайтом термінації, являє собою одиницю Т. — транскриптон. У прокаріотів після промотора розташована регуляторна ділянка — оператор, яка взаємодіє з білками — регуляторами Т. У транскриптоні еукаріотів роль такої регуляторної зони виконує акцепторна зона. У середині транскриптону (перед сайтом термінації) знаходяться структурні гени або цистрони. У прокаріотичних організмах первинний транскрипт часто містить РНК-копії одразу кількох генів, а в еукаріотів, як правило, — лише одиничного гена. В еукаріотів у кожному транскриптоні є неінформативна зона, з якою взаємодіють регуляторні транскрипційні фактори. Неінформативна частина в транскриптоні еукаріотів у середньому становить 90%, у прокаріотів — 10%. Біосинтез РНК здійснюється ДНК-залежними РНК-полімеразами. У прокаріотів існує тільки одна РНК-полімераза, яка каталізує синтез усіх трьох головних типів РНК, а в ядрах еукаріотів виявлені три РНК-полімерази: РНК-полімераза І, що синтезує попередник рРНК — пре-рРНК; РНК-полімераза ІІ, відповідальна за синтез пре-мРНК; РНК-полімераза ІІІ, що синтезує пре-тРНК і пре-5SРНК. РНК-полімерази складаються із декількох субодиниць — 2α, β, β′, що утворюють так званий кор-фермент, і σ-субодиниці, яка виконує регуляторну функцію і є одним із факторів ініціації Т. РНК-полімерази І, ІІ, ІІІ, які розпізнають різні промотори, містять різні за структурою субодиниці σ. Для Т. необхідні ДНК-матриця, фермент ДНК-залежна РНК-полімераза, 4 рибонуклеозидтрифосфати (АТФ, ГТФ, УТФ і ЦТФ) як попередники нуклеотидних елементів РНК, іони Mg2+, а також білкові фактори Т., які прискорюють або уповільнюють процес Т. Загальна схема РНК-полімеразної реакції може бути подана таким рівнянням:
n1АТФ + n2ГТФ + n3ЦТФ + n4УТФ → РНК + (n1 + n2 + n3 + n4)Н4Р2О7.
Процес Т. містить три стадії: ініціацію, елонгацію і термінацію. На першій стадії відбувається зв’язування РНК-полімерази з промотором ДНК-матриці за участю s-субодиниці ферменту, яка в подальшому дисоціює від кор-ферменту. Взаємодію РНК-полімерази з промотором полегшує ТАТА-фактор, який взаємодіє зі специфічною послідовністю нуклеотидів промотора — ТАТААА-(ТАТА-бокс). Останній змінює конформацію РНК-полімерази і забезпечує розкручування приблизно одного витка спіралі ДНК з утворенням транскрипційної вилки, в якій матричний ланцюг стає доступним для початку синтезу ланцюга РНК (рисунок).
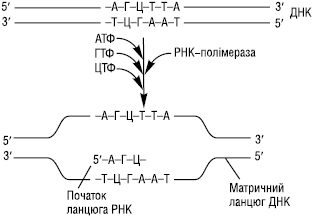
Рисунок. Т. РНК на матричному ланцюгу ДНК
Синтез РНК починається із включення в ланцюг першого (5′-кінцевого) нуклеотиду, яким у молекул РНК є АТФ або ГТФ. Після того як був синтезований олігонуклеотид із 8–10 нуклеотидних залишків, σ-субодиниця відділяється від РНК-полімерази, а замість неї до молекули ферменту приєднуються декілька факторів елонгації й починається наступна стадія. Синтез молекули РНК відбувається в напрямку 5′→3′ антипаралельно матричному ланцюгу ДНК. У процесі елонгації (подовження полінуклеотидного ланцюга) РНК-полімераза пересувається вздовж матриці ДНК і комплементарно їй послідовно приєднує нуклеотиди за допомогою фосфодіефірних зв’язків з утворенням зростаючого ланцюга РНК. Азотисті основи А, Г, Т, Ц ланцюга ДНК кодують включення в ланцюг, відповідно, основ У, Ц, А і Г. Унаслідок елонгації утворюється ДНК-РНК-гібрид, що складається з матричного ланцюга ДНК та РНК-транскрипту. Термінація Т. настає після досягнення РНК полімеразою сайтів термінації (стоп-сигналу). Вважають, що такими стоп-сигналами в транскриптоні є полі-(А)-послідовності, тому в ланцюгу РНК на 3′-кінці знаходяться комплементарні їм полі-(У)-послідовності. Цей сигнал розпізнається термінуючим білком — ρ-фактором. Взаємодія ρ-фактора з полімеразним комплексом приводить до дисоціації комплексу ДНК–фермент–РНК і вивільнення синтезованого ланцюга. Первинні транскрипти РНК або пре-РНК, що утворилися, є комплементарними копіями транскриптонів ДНК і містять як інформативні (екзони), так і неінформативні (інтрони) ділянки. Молекули пре-РНК піддаються далі посттранскрипційним змінам (процесингу), які містять ковалентні модифікації 3′- і 5′-кінців, вирізання інтронів, сплайсинг (зшивання) екзонів, нуклеозидні модифікації, внаслідок чого формуються зрілі функціональні молекули РНК.
Ряд антибіотиків гальмує Т. і використовуються для вивчення механізму цього процесу та як ЛП Актиноміцин Д вбудовується між сусідніми основами Г-Ц подвійної спіралі ДНК, що зумовлює локальну деформацію матриці ДНК і перешкоджає рухові РНК-полімерази по матриці. Він гальмує процес елонгації всіх типів РНК як у прокаріотів, так і у еукаріотів. Актиноміцин Д характеризується сильною протипухлинною та антибактеріальною дією, але внаслідок значної токсичності використовується частіше в біохімічних дослідженнях. Іншим важливим антибіотиком, що гальмує Т., є рифампіцин, який зв’язується з β-субодиницею РНК-полімерази прокаріотів і гальмує ініціацію синтезу РНК. На РНК-полімеразу еукаріотів рифампіцин не діє, тому широко використовується як антимікробний препарат. Він активний щодо мікобактерій туберкульозу, діє на стафілококи і менінгококи; рифампіцин використовують при запальних хворобах легень і дихальних шляхів (бронхіті, пневмонії), при остеомієліті та інших захворюваннях, викликаних чутливими до рифампіцину збудниками. Специфічну гальмівну дію на синтез РНК чинить α-аманітин — отруйна речовина із блідої поганки, який є сильним інгібітором РНК-полімерази ІІ еукаріотів і таким чином блокує у них синтез мРНК.
Биохимия / Под ред. Е.С. Северина. — М., 2003; Гонський Я.І., Максимчук Т.П. Біохімія людини. — Тернопіль, 2001; Губський Ю.І. Біологічна хімія. — К. — Тернопіль, 2000; Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека: В 2 т. — М., 1993. — Т. 2.