ТЕЛОМЕРАЗА (нуклеотидилтрансфераза) — фермент ембріональних та деяких інших клітин, що швидко діляться, забезпечує відновлення недореплікованих 5׳-кінців нових ланцюгів ДНК під час реплікації за рахунок синтезу нових теломер. Теломерними послідовностями, або просто теломерною ДНК, називають численні повтори (сотні або тисячі разів) олігонуклеотидів із шести залишків нуклеотидів у хребетних тварин (–GGGTTA–), із п’яти залишків нуклеотидів у комах (–GGTTA–) та семи залишків нуклеотидів у більшості рослин (–GGGTTTA–) на кожному кінці хромосоми. Наявність теломер необхідна для збереження генетичної інформації. Після завершення реплікації (подвоєння ДНК) хромосоми 5׳-кінці дочірніх ланцюгів ДНК лишаються недобудованими, тому що після видалення праймерів ці фрагменти залишаються недореплікованими. Це відбувається через нездатність ДНК-полімерази β, яка відповідає за заповнення ділянки після видалення праймерів, синтезувати ДНК від 3׳- до 5׳-кінця. Таким чином, під час кожного циклу реплікації 5׳-кінці синтезованих ланцюгів скорочуються. Проте ця втрата не зашкоджує генетичній інформації, бо відбувається за рахунок теломерів. Під час наступної реплікації 5׳-кінці знову залишаються недобудованими. Так, із кожним клітинним діленням ДНК хромосом послідовно скорочуються. Скорочення теломерів — важливий фактор, що визначає тривалість життя організму. Після приблизно 50 ділень клітини теломери вичерпуються, що є сигналом до апоптозу клітини. Відмирання клітин є одним із найважливіших механізмів старіння організму в цілому. Радянський учений О.М. Оловніков у 1971 р. висунув теорію, згідно з якою старіння розглядається як наслідок скорочення теломерних послідовностей. Згодом теорія була експериментально підтверджена та отримала назву теломеразної теорії старіння.
Т. відкрила у 1984 р. К. Грейдер, яка разом з Е. Блекберн та Дж. Шостак у 2009 р. стала Нобелівським лауреатом у галузі фізіології та медицини «За відкриття механізму захисту хромосом теломерами і ферментом теломеразою». До особливостей цього ферменту належить наявність простетичної групи молекули РНК (див. Рибонуклеїнові кислоти). Фрагмент РНК в активному центрі Т. є матрицею при синтезі теломерних повторів хромосом. За допомогою РНК Т. комплементарно (залишок аденіну з’єднується водневими зв’язками тільки з тиміном, залишок гуаніну — тільки з цитозином) прикріпляється до 3׳-кінця недосинтезованого дочірнього ланцюга ДНК. Фермент за принципом компліментарності послідовно подовжує 3׳-кінець ланцюга ДНК на один гексапептид –GGGTTA–. Синтез завжди відбувається від 5׳- до 3׳-кінця. Далі Т. пересувається вздовж ланцюга на один теломер та починає синтез нового фрагменту –GGGTTA–.
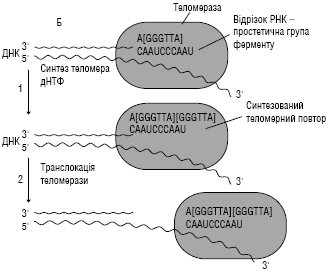
У більшості соматичних клітин Т. неактивна, тому що соматична клітина має довжину теломерної ДНК, достатню для часу життя клітини та її потомства. Проте невисоку активність Т. відзначають у клітинах з високою швидкістю оновлення: лімфоцитах, стовбурових клітинах кісткового мозку, клітинах епітелію, епідермісу шкіри та деяких інших. Наявність теломеразної активності в тих соматичних клітинах, де вона зазвичай не виявляється, може бути маркером злоякісної пухлини та індикатором несприятливого прогнозу.
Биохимия / Под ред. чл.-кор. РАН, проф. Е.С. Северина. — М., 2004; Nelson D.L., Cox M.M. Lehninger Principles of biochemistry. — New York, 2013.